Oxygen Transport by Hemoglobin and Myoglobin

Requires a Wolfram Notebook System
Interact on desktop, mobile and cloud with the free Wolfram Player or other Wolfram Language products.
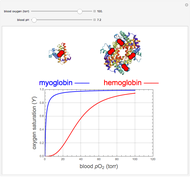
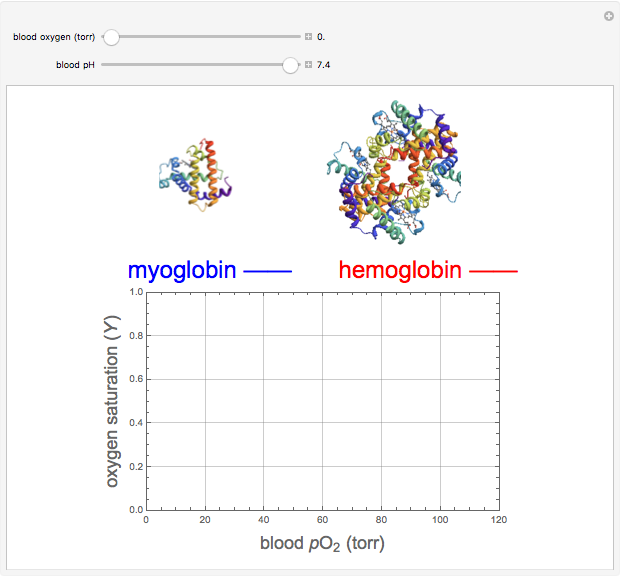
Hemoglobin, the essential component of red blood cells (erythrocytes), transports oxygen (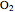 ) through the bloodstream from the lungs to all the tissues of the body. Hemoglobin also carries carbon dioxide (
) through the bloodstream from the lungs to all the tissues of the body. Hemoglobin also carries carbon dioxide ( ) back to the lungs to complete the process of respiration. Vertebrate hemoglobin is a nearly spherical protein molecule consisting of an array of four globin polypeptide chains, each containing a heme group, which is the oxygen-binding site. Its molecular weight is approximately 64,500 daltons. The oxygen uptake of hemoglobin exhibits cooperativity, such that each successively increases the affinity of the molecule for adding another , up to four. The saturation
) back to the lungs to complete the process of respiration. Vertebrate hemoglobin is a nearly spherical protein molecule consisting of an array of four globin polypeptide chains, each containing a heme group, which is the oxygen-binding site. Its molecular weight is approximately 64,500 daltons. The oxygen uptake of hemoglobin exhibits cooperativity, such that each successively increases the affinity of the molecule for adding another , up to four. The saturation  is a measure of the fractional occupancy of the oxygen-binding sites. It increases as a sigmoid-shaped function of the partial pressure of in its immediate environment. In the alveoli of the lungs,
is a measure of the fractional occupancy of the oxygen-binding sites. It increases as a sigmoid-shaped function of the partial pressure of in its immediate environment. In the alveoli of the lungs, 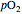 is approximately 100 torr.
is approximately 100 torr.
Contributed by: S. M. Blinder (April 2011)
Open content licensed under CC BY-NC-SA
Snapshots
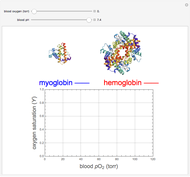
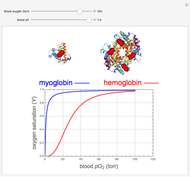
Details
The binding of oxygen to myoglobin is described by the equilibrium 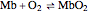 . The saturation can be defined by
. The saturation can be defined by  , where
, where  is the partial pressure for 50% saturation, about 1 torr for myoglobin. For hemoglobin, the equilibrium takes the form
is the partial pressure for 50% saturation, about 1 torr for myoglobin. For hemoglobin, the equilibrium takes the form 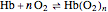 ,
,  to
to  . Here
. Here  , where
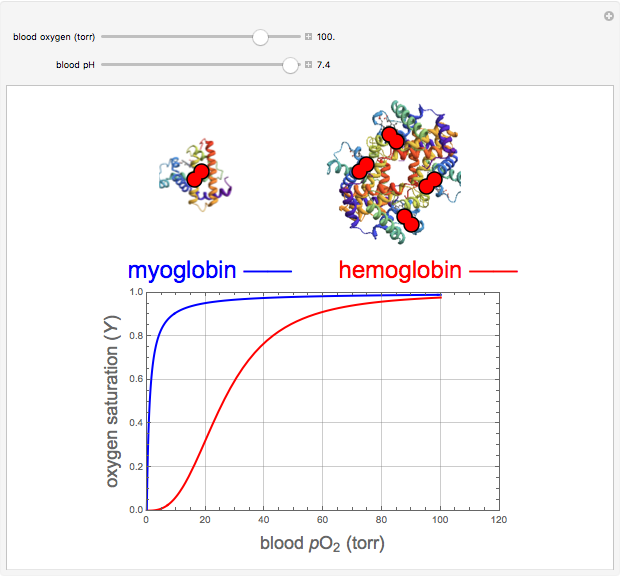
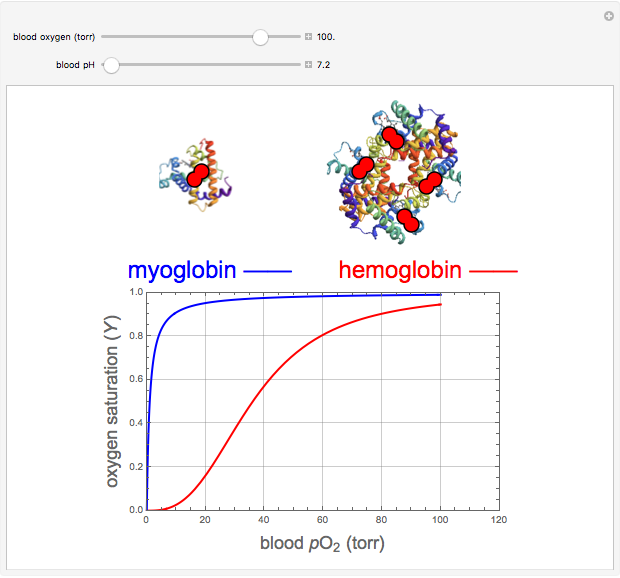
, where 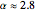 to account for the cooperativity of binding and
to account for the cooperativity of binding and 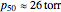 at pH 7.4 but slightly higher as the pH decreases.
at pH 7.4 but slightly higher as the pH decreases.
The central part of the heme group has this structure:  . The complexes with the iron atom.
. The complexes with the iron atom.
Reference: L. Stryer, Chap. 7, Molecular Design of Life, New York: W. H. Freeman, 1989.
Permanent Citation
 Effect of Altitude on Alveolar Oxygen Pressure
Effect of Altitude on Alveolar Oxygen Pressure
Natalie Gurevich and Kaitlyn Kuder Modeling Blood Glucose Levels
Modeling Blood Glucose Levels
Kevin Chen Ramachandran Analysis of Protein Backbone Dihedral Angles
Ramachandran Analysis of Protein Backbone Dihedral Angles
Daniel Barr Blood Donation Protocols
Blood Donation Protocols
D. Meliga and S. Z. Lavagnino Glycolysis
Glycolysis
Raymond Wang Light-Dependent Reactions in Photosynthesis
Light-Dependent Reactions in Photosynthesis
Derek Fu and Minha Kim Sensitivity Analysis of Transition Phases of Perturbed Gene Pathways with a Neural Network
Sensitivity Analysis of Transition Phases of Perturbed Gene Pathways with a Neural Network
Luca Zammataro DNA Base Pairing
DNA Base Pairing
S. M. Blinder 10-23 Deoxyribozyme Design Optimization
10-23 Deoxyribozyme Design Optimization
Brian M. Frezza Hodgkin-Huxley Action Potential Model
Hodgkin-Huxley Action Potential Model
Shimon Marom
-
 Ice Cube Melting in Water
Ice Cube Melting in Water
S. M. Blinder -
 Absorption Spectroscopy
Absorption Spectroscopy
S. M. Blinder -
 Height of Object from Angle of Elevation Using Tangent
Height of Object from Angle of Elevation Using Tangent
S. M. Blinder -
 Internal Rotation in Ethane and Substituted Analogs
Internal Rotation in Ethane and Substituted Analogs
S. M. Blinder -
 Topological Spaces on Three Points
Topological Spaces on Three Points
S. M. Blinder -
 Hydrogen Atom in Curved Space
Hydrogen Atom in Curved Space
S. M. Blinder -
 Multipurpose Tool
Multipurpose Tool
S. M. Blinder -
 Statistical Thermodynamics of Ideal Gases
Statistical Thermodynamics of Ideal Gases
S. M. Blinder -
 Bell's Theorem
Bell's Theorem
S. M. Blinder -
 Kepler's Mysterium Cosmographicum
Kepler's Mysterium Cosmographicum
S. M. Blinder -
 Bonding and Antibonding Molecular Orbitals
Bonding and Antibonding Molecular Orbitals
S. M. Blinder -
 Visible and Invisible Intersections in the Cartesian Plane
Visible and Invisible Intersections in the Cartesian Plane
S. M. Blinder -
 Heron's Formula
Heron's Formula
S. M. Blinder -
 How Zippers Work
How Zippers Work
S. M. Blinder -
 Mittag-Leffler Expansions of Meromorphic Functions
Mittag-Leffler Expansions of Meromorphic Functions
S. M. Blinder -
 Orbital Resonance in the Asteroid Belt
Orbital Resonance in the Asteroid Belt
S. M. Blinder -
 Jordan's Lemma Applied to the Evaluation of Some Infinite Integrals
Jordan's Lemma Applied to the Evaluation of Some Infinite Integrals
S. M. Blinder -
 Configuration Interaction for the Helium Isoelectronic Series
Configuration Interaction for the Helium Isoelectronic Series
S. M. Blinder -
 Structure and Bonding of Second-Row Hydrides
Structure and Bonding of Second-Row Hydrides
S. M. Blinder -
DNA Base Pairing
S. M. Blinder