Biodiversity in Spatial Rock-Paper-Scissors Games





Requires a Wolfram Notebook System
Interact on desktop, mobile and cloud with the free Wolfram Player or other Wolfram Language products.
The struggle for survival between competing bacteria species in a petri dish can create beautiful spiral patterns—until one type conquers all the available space, destroying the entangled structure. Kerr et al. made these observations when they mixed three E. coli populations exhibiting cyclic dominance (Kerr et al., 2002): A beats B beats C beats A. As shown in (Reichenbach et al., 2007; 2008), the mobility of the bacteria, that is, their spatial relocation rate, is crucial for the stability of coexistence. Using this Demonstration, you can simulate a spatial rock-paper-scissors game and observe the role mobility plays.
Contributed by: Martin Weidl (March 2011)
Suggested by: Erwin Frey
After work by: Tobias Reichenbach, Mauro Mobilia, and Erwin Frey
Open content licensed under CC BY-NC-SA
Snapshots
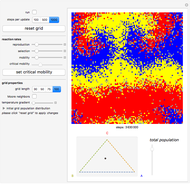
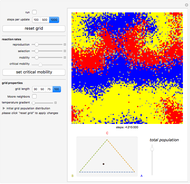
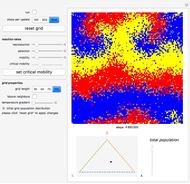
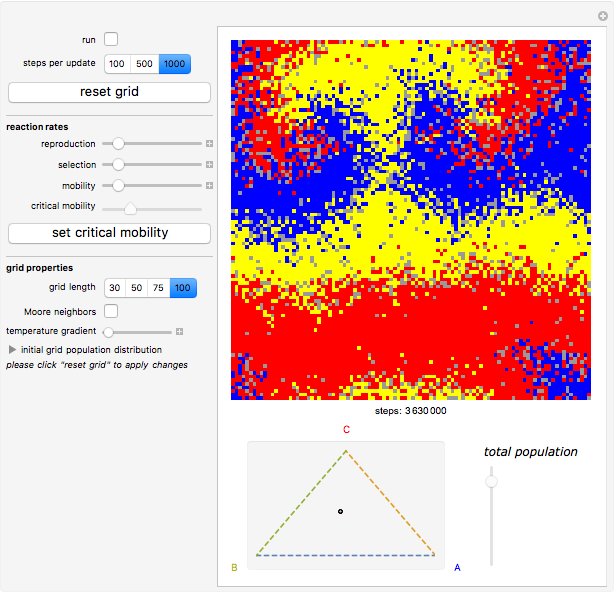
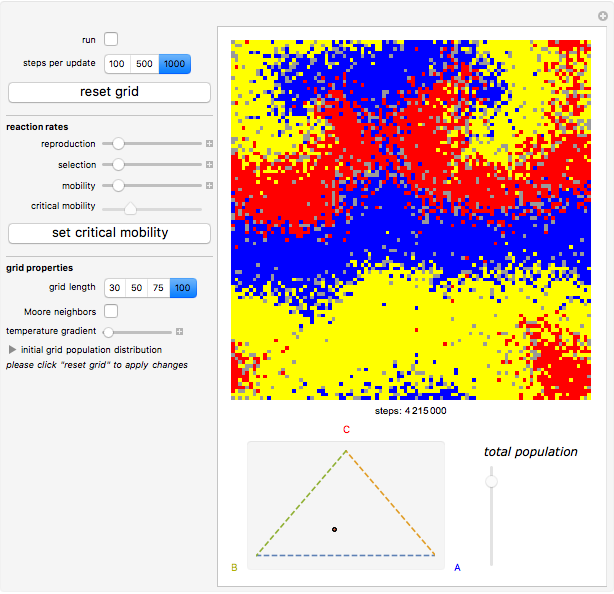
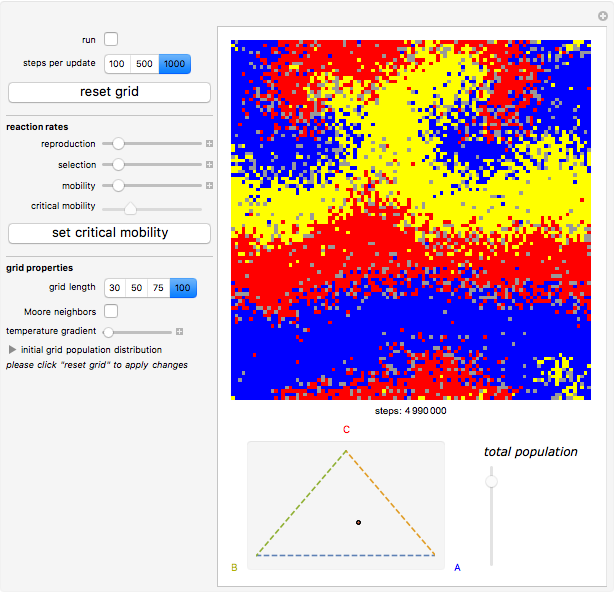
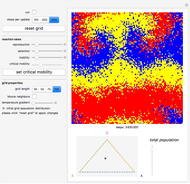
Details
Random cells on a square lattice with periodic boundary conditions are populated with three species, A (blue), B (yellow), and C (red) that cyclically dominate each other, as in a game of rock-paper-scissors. The simplex below the grid shows you the relative distribution of the species. When the simulation starts, three different reaction types occur at rates that you can adjust with the corresponding sliders:
1. In a reproduction process, a pair of adjacent cells is randomly chosen. If either cell is currently empty, it is populated with the species that occupies the second cell. If the second cell is empty as well, both are left unoccupied.
2. A selection process only occurs if the two cells of a random pair of nearest neighbors are occupied by different species. Then, the cell that is populated with the weaker species (in the sense of the rock-paper-scissors principle described above) is vacated.
3. Whenever a mobility process occurs, two random nearest-neighbor cells simply exchange their respective contents.
If the frequency of the last reaction is low enough, the cells occupied by each of the species, which continuously chase each other, form a spiral-like pattern slowly crawling across the grid. With increasing mobility rate, the size of the spirals grows and the structure becomes more and more coarse-grained, until it eventually dissolves completely as one species is able to conquer the entire grid. This disappearance of biodiversity is inevitable for any setting of the mobility slider. Beyond a critical mobility rate, however, the average duration of coexistence grows slower than linearly in the size of the grid and you will notice that a one-species state is reached much earlier.
The population frequencies of the three species are visualized as a point on the triangular simplex below the grid: close proximity to one of its vertices signals that a large share of the total population belongs to the species indicated next to the vertex. The fraction of grid cells occupied by any of the three species is shown by the slider next to the simplex.
You can see the finer details of the spirals more easily on larger grids, but the formation of any discernible structure takes longer as well. You can also examine the effects of choosing pairs of Moore neighbors (i.e., two cells sharing an edge or a corner) instead of the default von Neumann neighbors (i.e., two cells with a common edge), or try setting a temperature gradient so that reactions occur more often near the center of the grid.
Finally, you can adjust the probability distribution that determines the initial grid population and even include blocked cells, which do not take part in any interaction, to study how the game evolves in less regular topologies.
References: B. Kerr, M. A. Riley, M. W. Feldman, and B. J. M. Bohannan, "Local Dispersal Promotes Biodiversity in a Real-Life Game of Rock-Paper-Scissors," Nature, 418, 2002 pp. 171–174. T. Reichenbach, M. Mobilia, and E. Frey, "Mobility Promotes and Jeopardizes Biodiversity in Rock-Paper-Scissors Games," Nature, 448, 2007 pp. 1046–1049. T. Reichenbach, M. Mobilia, and E. Frey, "Self-Organization of Mobile Populations in Cyclic Competition," J. Theor. Biol., 254(2), 2008 pp. 368–383.
Permanent Citation
"Biodiversity in Spatial Rock-Paper-Scissors Games"
http://demonstrations.wolfram.com/BiodiversityInSpatialRockPaperScissorsGames/
Wolfram Demonstrations Project
Published: March 7 2011
 Stochastic Rock-Paper-Scissors Population Dynamics
Stochastic Rock-Paper-Scissors Population Dynamics
Martin Weidl Ecosystems with Ghost Invasive Species
Ecosystems with Ghost Invasive Species
Wilfried Gabriel and Maria Stockenreiter Predator-Prey Ecosystem: A Real-Time Agent-Based Simulation
Predator-Prey Ecosystem: A Real-Time Agent-Based Simulation
Hiroki Sayama Garbage Collection by Ants
Garbage Collection by Ants
Hiroki Sayama Bifurcation in a Model of Spruce Budworm Populations
Bifurcation in a Model of Spruce Budworm Populations
Selwyn Hollis Ecology of Reef Systems
Ecology of Reef Systems
Cameron Berglund Genetic Drift of Allele Frequency
Genetic Drift of Allele Frequency
John Hawks Pathogen Spillover Effects on Coexistence of Competing Host Species
Pathogen Spillover Effects on Coexistence of Competing Host Species
Erin A. Mordecai Stochastic Model of Seed and Spore Germination
Stochastic Model of Seed and Spore Germination
Mark D. Normand and Micha Peleg Quasi-Rent in Open Access Fisheries
Quasi-Rent in Open Access Fisheries
Arne Eide